


")





Antiguamente el concepto de la producción sobre la carne porcina hace referencia a una producción en condiciones de poca higiene, contenido de alto tenor de grasa y colesterol. Entretanto, hoy la carne porcina es el resultado de la evolución basada en la selección genética, nutrición, manejo y estatus sanitario de los rebaños, presentando tenores reducidos de grasas y calorías y mayor porcentaje de carne magra en relación a 25 años atrás (Figura 1).
A lo largo de los últimos 25 años se han producido distintos cambios en la industria porcina. Estos cambios contribuirán a la mejoría fenotípica presentada por los cerdos en el peso de faena (110 a 122 kg), rendimiento de carcasa (71 a 74%), y no mejorías en la cualidad y cantidad de carne depositada en la carcasa para las razas modernas de Yorkshire, Duroc, Hampshire, Pietran e Landrace, las que fueron utilizadas para los programas genéticos de producción de cerdos comerciales actuales.
Los programas de alimentación también sufrieron cambios en los últimos 25 años, permitiendo aumentar de manera significativa el grado de tejido magro. Dentro de los principales cambios nutricionales que más impactan sobre el desempeño de los cerdos, se puede citar:
• Peletización de la ración, permitió aumentar la ganancia de peso diaria reduciendo el consumo y mejorando la conversión alimentaria (Wondra et al., 1995);
• Inclusión de antibióticos y promotores de crecimiento en las dietas, promovió mejores índices de crecimiento y mejor eficiencia alimentaria (Cromwell et al., 1996);
• Adición de grasa a las dietas: mejoró la eficiencia alimentaria (De la Llata et al., 2001);
• Nutrición basada en el concepto de proteína ideal, permitió mantener la producción ideal de los aminoácidos esenciales con lisina (primer aminoácido limitante para la deposición de tejido magro), promoviendo mejoras en la eficiencia alimentaria, en el crecimiento en la ganancia muscular, y una reducción de los niveles proteicos de las raciones (Robinson et al., 2000; Witte et al., 2000);
• Un mejor entendimiento de los requerimientos nutricionales para las fases específicas de la vida del cerdo, permitiendo explorar mejor las curvas de crecimiento y de deposición de proteína.
En una revisión sobre la producción de calor de los terminadores modernos, Brown-Brandl et al. (2004), concluían que los genotipos modernos presentaban un aumento de producción de calor en un 18.1% (aproximadamente 1% al año) de 1984 a 2002. Podemos atribuir a este aumento, cambios en las composiciones corporales de los cerdos actuales (menos grasa y más músculo; Figura 2 y 3) y no al aumento de las tazas de “turnover” proteico. De acuerdo con Ball et al. (2008), para cada 2,8% de aumento en el porcentual de tejido magro, una producción de calor aumenta en torno de 18.7%. El aumento presenta un impacto significativo sobre las instalaciones y manejos adoptados para estos animales. El aumento de producción de calor endógeno debe ser considerado, principalmente en regiones con clima tropical, en el momento en que está siendo hecha la construcción de las instalaciones y durante las formulaciones de las dietas, Estos cambios ocurridos en la composición corporal de los cerdos y en el aumento de las tazas de “turnover” proteico, también contribuyen para mayores necesidades de energía y aminoácidos.
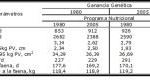
En la tabla 1, se puede observar como los cambios en el potencial genético y en el programa nutricional alteraron el potencial productivo de los cerdos en la fase de terminación.
Tabla 1. Efecto de la ganancia genética y del programa nutricional sobre el desempeño de los cerdos en la fase de terminación.
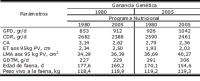
GPD = ganancia de peso diario; CDR = consumo de ración diaria; CA = conversión alimentaria; ET = espesura de tocino mensurado en la 10a costilla; LMA = área de músculo Longissimus dorsi mensurado en la 10a costilla; GDTM = ganancia diaria de tejido magro; PV = peso vivo.
(Fuente: adaptado de Fix et al., 2010).
Con base en los datos presentados, se puede inferir que una reducción en 15% de los días de faena y un 45% de aumento en la eficiencia de carne magra pueden ser atribuidos casi en una misma proporción que el mejoramiento genético y los avances de la nutrición. Una mejora del 33% en la conversión alimentaria representa 2/3 dos resultados de cambios en el programa nutricional. La reducción de la espesura de tocino es atribuida de manera exclusiva al proceso de selección genética, en cuanto que el porcentual de tejido magro parece estar relacionado de manera igual a la genética y al programa nutricional.
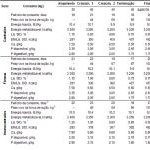
En la tabla 2 se puede visualizar la recomendación nutricional dada por TOPIGS (progenie con el macho Tempo, 2010) para las fases de crecimiento y terminación. Esta recomendación busca maximizar no solo el potencial de deposición proteico, sino también permite explorar de forma más completa los potenciales de los diferentes sexos y sus respectivas fases de crecimiento, permitiendo así, optimizar el potencial del genotipo.
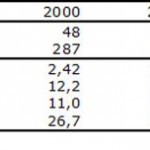
El intenso trabajo de mejoramiento genético realizado en los cerdos terminadores, con el objetivo de obtener una mayor eficiencia alimentaria y carcasas con mayor tenor de carne magra, también tienen elevado las alteraciones significativas en las matrices actualmente disponibles en el mercado. Las matrices modernas son más precoces, más productivas y poseen mayor peso corporal, por lo tanto, siendo nutricionalmente más exigentes. La productividad de la hembra aumento de forma substancial en los últimos anos, dado a el manejo y los avances en la selección genética, selección basada en parámetros como: tamaño de camada, intervalo desmame estro y eficiencia en la lactación. Es posible observar en la Tabla 3 una indicación del tipo de progreso alcanzado en las 10% mejores granjas comerciales hoy en Holanda.
Tabla 2. Programa alimentario sugerido y requerimientos nutricionales de las dietas utilizadas para progenie de TEMPO (Fuente: adaptado del manual de alimentación de la progenie Tempo, TOPIGS do Brasil 2010)

1 Lisina digestivel ileal padronizada
2 Para animais imunocastrados a troca da Ração Terminação para a Final deverá ser realizada ao final da primeira semana imediatamente após a aplicação da 2ª dose da vacina auto-imune (Vivax®).
3 Consumo estimado na fase, respectivamente para Castrados, Fêmeas e Imunocastrados.
Tabla 3. Progreso alcanzado en la productividad en las 10% mejores granjas comerciales de Holanda entre 2000 e 2009 (Fuente: TOPIGS Netherlands Annual Report 2010)

Los datos presentados en la Tabla 3 demostrando la eficiencia de selección genética balanceada, que trae asociada una mayor capacidad reproductiva de las hembras de mayor vigor y supervivencia de los lechones, permitiendo la producción de camadas sin aumentar la mortalidad de los lechones (Figura 4). Los resultados indican un crecimiento estable de 0,35 lechones desmamados/cerda/año. Estos valores nos permiten una estimación para 2020 de 15 – 16 lechones nascidos vivos/camada o 33 lechones desmamados/cerda/año. En este momento no existen evidencias de que este mejoramiento haya alcanzado su límite máximo.
Dado a que el avance genético hace a las cerdas más productivas, las mismas se vuelven más demandantes y menos resistentes a los desafíos nutricionales. Las necesidades nutricionales de las cerdas modernas, y la disponibilidad de nutrientes de la dieta para las mismas, son poco conocidas en comparación a lo que se sabe para cerdos en fase de crecimiento o terminación.
Las estimaciones de energía para cerdas gestantes fueron determinadas por Noblet et al. (1997), en donde los autores afirman que más del 60% de las exigencias de energía de las cerdas gestantes son representadas por la mantenencia, estando en torno de 0,44 MJ (o 105 kcal) de EM/ kg de peso metabólico (PC0.75)/ día, en condiciones de termo-neutralidad. Este valor, según los mismos autores no están significativamente influenciados por el orden de parto, gestación y fase da gestación. Más recientemente, Samuel et al. (2007) revalidan las necesidades de energía metabolisable de manteniencia para genotipos modernos (alta taza de deposición de tejido magro e alta prolificidad) y observaron una necesidad de 0,50 MJ (o 120 kcal) de EM/ kg de peso metabólico (PC0.75)/ día. Los mismos autores concluyeron que el valor sugerido por Noblet et al. (1997) e NRC (1998) para la actual población de cerdas esta debajo de las necesidades reales en aproximadamente 14%. Una probable explicación para los cambios en estas necesidades estan relacionados a una disminución en el porcentaje de grasa corporal y a un aumento del porcentaje de masa proteica, así como al aumento de las tazas de “turnover” proteico de los genotipos modernos.
Durante la gestación, las exigencias proteicas de mantenimiento han sido estimadas con gran variabilidad, con valores entre 50 a 133 g de proteína/día (Patience, 1996), pudiendo estar relacionada la masa proteica corporal. Fuller et al., (1989) e NRC (1998) estimaron una exigencia diaria de lisina para mantenimiento en 36 mg para cada kg de peso metabólico (PC0,75). Recientemente, Samuel et al. (2008a) validaron estas necesidades de lisina para mantenimiento de cerdas de genotipo moderno usando la oxidación de aminoácidos como indicadores. Estos autores encontraron que la necesidad de lisina es de 49 mg/ kg de peso metabólico (PC0,75); este valor excede a la actual recomendación (Fuller et al., 1989; y NRC 1998) en 30%.
Existen más informaciones disponibles sobre las exigencias de lisina de que de otros aminoácidos esenciales. Entretanto, si aplicamos la relación de proteína ideal utilizando la necesidad de lisina propuesto por Samuel et al. (2008a), la necesidad de metionina sería 40% superior a la recomendación actual. La metionina desempeña una función esencial durante la gestación, incluyendo la metilación de DNA durante el desenvolvimiento embrionario/fetal, lo que la torna extremamente importante para la regulación de la expresión genética. Dourmad y Etienne (2002) concluyeron que la necesidad de treonina durante a gestación para las cerdas modernas es mayor que el valor propuesto por NRC (1998), estos autores atribuyen esta diferencia a una mayor retención de nitrógeno diaria observada en las cerdas modernas durante a gestación. Estos resultados corroboran la hipótesis de que las necesidades de aminoácidos de los genotipos modernos serán mayores, en virtud de una mayor capacidad de deposición de tejido magro y de “turnover” proteico. Analizando estudios recientes con cerdas de genotipos modernos, ha sido dada una atención particular al crecimiento fetal (McPherson et al., 2004), desenvolvimiento de las glándulas mamarias (Ji et al., 2006), y a el crecimiento materno (Ji et al., 2005). Los resultados obtenidos por estos autores indican un crecimiento cúbico, tanto de tejido mamario, cuanto de los fetos, principalmente, a partir de los 70 días de gestación. Resultados estos, superiores a los observados en estudios similar es en las décadas de 80 y 90.
Con base en los resultados de las recientes investigaciones deben ser estimulado el establecimiento de programas nutricionales basados en más de una fase, y no más en una única dieta durante todo o período de gestación para cerdas nulíparas (McPherson et al., 2004) (Figura 5 e 6).
Informaciones actualizadas sobre la necesidad de energía para matrices lactantes modernas son limitadas. El consumo inadecuado de energía durante la lactación hace que la matriz movilice nutrientes de diferentes tejidos corporales, llevando a una significativa pérdida de peso. La mayoría de los trabajos afrontan las exigencias energéticas de mantenencia semejantes entre cerdas gestantes y lactantes. Noblet et al. (1990) sugiere diferentes valores, 0,46 (o 110 kcal) y 0,44 (o 105 kcal) MJ de EM/ kg de peso metabólico (PC0,75) para lactación y gestación respectivamente. Estas diferencias parecen ser relativamente pequeñas, del orden de 5%. Más recientemente, Samuel et al. (2007a, b, c, d) reestimaron la energía de mantenencia para cerdas lactantes en 0,51 (o 122 kcal) ± 0,07 (o 16,73 kcal) MJ de EM/ kg de peso metabólico (PC0,75), 10% superior al valor propuesto por Noblet et al. (1990).
En sistemas de manejo convencionales, lechones recién nacidos son capaces de atingir solamente, una fracción de su real potencial de crecimiento durante la fase de lactación (Harrel et al., 1993). En los últimos años, esto ha llevado investigadores a focalizar en la importancia de los nutrientes y en particular de los aminoácidos, para optimizar el potencial de las glándulas mamarias durante la lactación (Kim e Wu, 2008). El crecimiento da glándula mamaria durante esta fase afecta la cantidad de leche producida por las cerdas, y en consecuencia el crecimiento de los lechones (Kim et al., 2000a). De esta forma, el manejo nutricional adoptado durante a lactación deberá priorizar el máximo crecimiento mamario. Cuando las cerdas no reciben cantidades adecuadas de aminoácidos en la dieta, proteínas del tejido materno, particularmente, de las proteínas musculares esqueléticas, son movilizadas para atender a la producción de leche. La movilización excesiva de proteína materna, generalmente, resulta en fallas en la actividad reproductiva subsecuente (Jones e Stahly, 1999).
El cerdo moderno presenta considerable potencial genético de ganancia y eficiencia de conversión alimentaria, mientras tanto, para establecer un adecuado programa de nutrición para estos animales, se debe considerar el material genético de la granja, las necesidades nutricionales de estos animales, los factores que afectan esas necesidades, y se debe poseer entendimiento de los diversos aspectos metabólicos de la interacción entre el genotipo, nutrición y sanidad. Este entendimiento es fundamental para que se pueda alcanzar, al mismo tiempo, la máxima productividad, longevidad y rentabilidad. Así siendo, las dietas den ser formuladas y ajustadas de acuerdo con el genotipo de cada fase de la vida del animal, procurando siempre optimizar el potencial de consumo y mantener el sistema inmunológico equilibrado.
Literatura Citada
Ball, R.O.; R.S. Samuel and S. Moehn. 2008. Nutrient requirents of prolific sows. Advances in Pork Production. 19: 223.
Brown-Brandl, T. M., J. A. Nienaber, H. Xin, & R. S. Gates. 2004. A literature review of swine heat production. Trans. ASAE. 47(1): 259-270.
Chen, P., Baas, T.J., Mabry, J.W., Dekkers, J.C.M., Koehler, K.J., 2002. Genetic parameters and trends for lean growth rate and its components in U.S. Yorkshire, Duroc, Hampshire, and Landrace pigs. J. Anim. Sci. 80, 2062–2070
Canadian Centre for Swine Improvement. 2007. Annual report 2006/2007. http://www.ccsi.ca/meetings/annual/ann2007.pdf/
Cromwell, G.L., Davis, G.W., Morrow, W.E.M., Primo, R.A., Rozeboom, D.W., Sims, M.D., Sanisiewski, E.P., Ho, C.H., 1996. Efficacy of antimicrobial compound U-82, 127 as a growth promoter for grow-finishing pigs. J. Anim. Sci. 74, 1284–1287.
De la Llata, M., Dritz, S.S., Tokach, M.D., Goodband, R.D., Nelssen, J.L., Loughin, T.M., 2001. Effects of dietary fat on growth performance and carcass characteristics of growing–finishing pigs reared in a commercial environment. J. Anim. Sci. 79, 2643–2650.
Dourmad, J.Y., & M. Etienne. 2002. Dietary lysine and threonine requirements of the pregnant sow estimated by nitrogen balance. J. Anim. Sci. 80: 2144-2150.
Fix, J.S., Cassady, J.P., E.van Heugten, D.J. Hanson, M.T. See. 2010. Differences in lean growth performance of pigs sampled from 1980 and 2005 commercial swine fed 1980 and 2005 representative feeding programs. Liv. Sci. 128: 108-114
Fuller, M.F.; McWilliams, R.; Wang, T.C. & Giles, L.R. 1989. The optimum dietary aminoacid pattern of growing pigs. 2. Requirements for maintenance and for tissue protein accretion. British J. Nutr., 62:225-267.
Harrell, R. J., M. J. Thomas, and R. D. Boyd. 1993. Limitations of sow milk yield on baby pig growth. Pages 156–164 in Proc. Cornell Nutr. Conf. Cornell Univ., Ithaca, NY.
Jones, D. B., and T. S. Stahly. 1999. Impact of amino acid nutrition during lactation on body nutrient mobilization and milk nutrient output in primiparous sows. J. Anim. Sci. 77:1513–1522.
Ji, F., W. L. Hurley, and S. W. Kim. 2006. Characterization of mammary gland development in pregnant gilts. J. Anim. Sci. 84:579–587.
Ji, F.; Wu, G.; Blanton, J.R. and Kim, S.W. 2005. Changes in weight and composition in various tissues of pregnant gilts and their nutritional implications. J. Anim. Sci., 83:366-375.
Kim, S. W., and G. Wu. 2008. Regulatory role for amino acids in mammary gland growth and milk synthesis. Amino Acids doi:10.1007/s00726-008-0151-5.
Kim, S. W., W. L. Hurley, I. K. Han, and R. A. Easter. 2000a. Growth of nursing pigs related to the characteristics of nursed mammary glands. J. Anim. Sci. 78:1313–1318.
McPherson, R.L.; Ji, F.; Wu, G.; Blanton JR. J.R. and Kim, S.W. 2004. Growth and composition changes of fetal tissues in pigs. J. Anim. Sci., 82:2534-2540.
Noblet, J.; Dourmad, J.Y.; Etienne, M. & Le Dividich, J. 1997. Energy metabolism in pregnant sows and newborn pigs. J. Anim. Sci., 75:2708-2714.
Noblet, J.; Dourmad, J.Y. and Etienne, M. 1990. Energy utilization in pregnant and lactating sows: modeling of energy requirements. J. Anim. Sci., 68:562-572.
NATIONAL RESEARCH COUNCIL. – NRC. Nutrient requirements of swine. 10 ed. Washington, D.C. : National Academic Science, Committee Animal Nutrition. Subcommittee of Swine Nutrition, 1998. 189 p.
Patience, J.F. 1996. Meeting the energy and protein requirements of the high producing sow. Anim. Feed Sci. Technology, 58:49-64
Robison, O.W., Christian, L.L., Goodwin, R., Johnson, R.K., Mabry, J.W., Miller, R.K., Tokach, M.D., 2000. Effects of genetic type and protein levels on growth of swine. Proc. of the ASAS Meetings, Indianapolis, IN, pp. 1–9.
Samuel, R. S.; S. Moehn; P. B. Pencharz; and R. O. Ball. 2008a. Dietary lysine requirement for maintenance is 49 mg/kg0.75 in a population of modern, high producing sows. Advances in Pork Production. 19: A1.
Samuel, R.S.; S. Moehn; P.B. Pencharz; and R.O. Ball. 2007. Estimates of energy requirements during gestation and lactation in sows. Advances in Pork Production. 18: A9.
Topigs do Brasil. Manual de alimentação da progênie TEMPO 2010.
Witte, D.P., Ellis, M., McKeith, F.K., Wilson, E.R., 2000. Effect of dietary lysine level and environmental temperature during the finishing phase on the intramuscular fat content of pork. J. Anim. Sci. 78, 1272–1276.
Wondra, K.J., Hancock, J.D., Behnke, K.C., Hines, R.H., Stark, C.R.Stark, 1995. Effects of particle size and pelleting on growth performance, nutrient digestibility and stomach morphology in finishing swine. J. Anim. Sci. 73, 757–763.
Please follow and like us: